Dilleniaceae
Dilleniaceae Salisb., Parad. Lond. 2, 1: ad t. 73 (1807), nom. cons. (‘Dilleneae’).
J.W. Horn
Trees, shrubs, or lianas, rarely subshrubs or rhizomatous herbs; vestiture of sclerified and/or silicified simple and sometimes also fasciculate trichomes; glandular trichomes very rare. Leaves spirally arranged, very rarely opposite; blades petiolate or uncommonly sessile, simple, or rarely threefold pinnatisect to pinnately compound; margins entire or toothed; venation craspedromous, semicraspedromous, brochidodromous, or eucamptodromous, frequently with ± straight, parallel secondaries terminating in the teeth (when present), and rigidly percurrent tertiaries; stipules 0, but the petiole sometimes with persistent or caducous amplexicaul wings, and often with a broad insertion. Plants synoecious, or rarely structurally androdioecious and functionally dioecious. Inflorescences terminal, axillary, or ramiflorous, determinate; frequently a thyrsoid with cincinnate or modified dichasial partial inflorescences, a panicle, or monad, sometimes a botryoid or cincinnus; pedicels commonly with apical articulation. Flowers small to very large, actinomorphic or (mainly in the androecium) monosymmetric, hypogynous or very rarely partly epigynous, without nectar; receptacle flat or infrequently conical; sepals (3)4–5(–18), equal to unequal, typically free, membranaceous to coriaceous, imbricate (quincuncial when 5), always persistent, slightly to substantially accrescent in fruit; petals (2)3–5(–7), free, elliptic to obovate, often emarginate, typically white or yellow, frequently crumpled in bud, imbricate (quincuncial when 5), typically caducous; stamens (1 or 3–)5–400(–900), occasionally partly staminodial, typically marcescent, free or sometimes the filaments basally to nearly fully connate and then typically grouped into 1, 2, 3, or 5 fascicle(s), rarely forming a short tube; anthers basifixed, dithecal and tetrasporangiate, linear to oblong to subglobose, dehiscing via longitudinal slits, apical clefts, or apical pores; connective sometimes thickened, distinctly separating the thecae, and occasionally protruding apically as a short mucro; gynoecium apocarpous to, less frequently, hemisyncarpous, of 1–10(–20) carpels arranged in1whorl (very rarely 2 whorls); stylodia free; stigmas punctiform,minute, and not differentiated in shape from the stylodia, or the stigmas peltate with an annuliformor, infrequently, irregular margin; ovules 1–80, anatropous to campylotropous,when 1, apotropous,when 1–2, 1 apotropous and 1 epitropous, erect, or when 4 or more, pleurotropous and syntropous, bitegmic, crassinucellate; placentation submarginal, in 2(4, 6) vertical rows, or basal when ovules 1–2. Fruit most frequently a follicle or aggregate of follicles (sometimes basally coherent), or indehiscent and enclosed by the fleshy, accrescent sepals, less often a fleshy capsule, berry, or aggregate of nutlets; aril fleshy to scarious and oily or waxy, funicular, rarely vestigial; seed coat with typically heavily sclerotized or sometimes cutinized endotesta; raphe short; endosperm fleshy, oily or sometimes also starchy, abundant; embryo straight, minute.
A pantropical family with a largely Gondwanic distribution, extending into temperate Australia, containing 10 genera and c. 500 species.
Characters of Rare Occurrence. Rhizomatous herbs in Acrotrema. Subshrubs mostly or only with cataphylls and always with green, photosynthetic, sympodial, aerial stems that are terete and ephedroid to flattened and phyllocladous (sometimes dimorphic) in Hibbertia subg. Pachynema. Lignotuberous shrubs in several species of Hibbertia and Tetracera masuiana. Stilt roots occasionally
present in several Dillenia spp. and consistently present in D. borneensis, D. grandifolia, and D. reticulata (Hoogland 1952, 1959). Leaves pinnatisect to pinnate in a fewSri Lankan Acrotrema spp., sessile and rarely amplexicaul to entirely perfoliate (some Hibbertia). Leaves opposite in Doliocarpus
pruskii and Hibbertia coriacea and with persistent (Davilla spp., Didesmandra, Dillenia spp., Schumacheria) or caducous (Acrotrema, Dillenia spp.) amplexicaul wings. Leaf domatia in Doliocarpus spp. and Tetracera spp. Leaves turning yellow or golden brown during extreme drought and regreening after rain (diallagy; George 2002) in Hibbertia hypericoides and H. spicata subsp. leptotheca (and perhaps other Hibbertia spp. in regions with arid climates). Plants structurally androdioecious and functionally dioecious in neotropical Tetracera spp. Petals absent in Dillenia celebica, D. grandifolia, and D. serrata (Hoogland 1952, 1959). Corolla orange in Hibbertia comptonii, H. miniata, and H. stellaris, pink to deep red in several species of Hibbertia subg. Pachynema, and deep red in Dillenia pteropoda, inconsistently deep red in a few Dillenia or pink in a few Asian Tetracera (Hoogland 1953). Corolla not spreading at anthesis in Dillenia papuana and several putatively relatedDillenia. Fertile androecium distinctly heterantherous in Didesmandra, many Dillenia, Hibbertia heterotricha, H. margaretae, H. nana, H. pulchella. Stamens fewer than 5 in Hibbertia hirsuta (1–3), some populations of H. fasciculata and H. racemosa(3), Hibbertia (Pachynema) praestans (4), and H. rufa (4). Floral receptacle conical in the region of the gynoecium in Dillenia and Hibbertia baudoinii. Carpels more than 5 in most Dillenia spp., Hibbertia grossulariifolia, and some populations of H. scandens.
Vegetative Morphology. The family varies from tall, sometimes buttressed trees (Dillenia spp.), small to medium-sized, tortuous trees (Curatella), small to medium-sized rosette trees (New Caledonian Hibbertia), to shrubs (Hibbertia; some Davilla, Doliocarpus, and Tetracera), lianas (e.g., Davilla, Doliocarpus, Neodillenia, Pinzona, Tetracera, some Hibbertia), subshrubs with ephedroid or phyllocladous, photosynthetic aerial shoots and foliage typically of cataphylls (Hibbertia subg. Pachynema), and perennial herbs with a woody rhizome (Acrotrema). The bark often has a characteristic rich red- or orange-brown color and abundantly exfoliates in thin, papery plates, flakes, or strips, or the outer bark is gray, and exfoliates in flakes to reveal a rich-brown younger bark. Scandent species climb by twining, and the extension growth of large, climbing individuals of Delimoideae and Doliocarpoideae is often conspicuously curved or hooked.
Virtually the entire aerial plant body of Hibbertia subg. Pachynema, which consists of photosynthetic stems with cataphylls, represents inflorescences, as is shown by the occasional production of ‘normal’ foliage leaves at the basalmost nodes of a developing shoot in a few species (Craven and Dunlop 1992): the shoot systems can be compared with a compound thyrsoid or panicle. Foliage of some early branches of, e.g., Hibbertia subg. Pachynema, are mostly the prophylls of singleflowered sympodia, and the leaves of Hibbertioideae may represent heterotopic inflorescence bracts. As in some Dillenioideae, the β-prophyll is displaced onto the branch borne in its axil. Sympodia of other species of H. subg. Pachynema have several cataphyll-bearing nodes between the base of the branch and the flower. Shoot systems in Hibbertia subg. Adrastaea are fundamentally similar, though they do not show metatopic displacement of the prophylls. In Dillenia spp., axillary buds are sometimes accompanied by two serial buds that are displaced (by concaulescence), sometimes a considerable distance up the stem.
The foliage is typically evergreen, though in some Dillenia indigenous to parts of monsoonal SoutheastAsia, it is deciduous.Young leaves exhibit conduplicate or plicate-conduplicate ptyxis. The petioles are usually distinct, and oftenhave a broad, adaxial groove or channel. The bases of the petiolar wings completely ensheath the stem in Acrotrema, Davilla alata and allied species, Didesmandra, Dillenia spp., and Schumacheria.These petiolarwings, which may be persistent or caducous, enclose the developing terminal bud; they do not receive an independent trace from the cauline stele, and are not stipular (Dickison 1969).
Inall subfamilies exceptHibbertioideae, higher orders of leaf venation are typically very regularly organized; rigidly percurrent, scalariform tertiary venation and well-developed areoles, oriented with the lower orders of venation, are common. Leaf venation in Hibbertioideae is distinctly less organized; percurrent tertiaries are rare and never rigidly scalariform, and areolation is typically incomplete or lacking (Rury and Dickison 1977). Ericoid leaves are characteristic of many species of Hibbertia subg. Hemistema and the H. hemignosta complex (sect.Candollea, subg. Hibbertia:Wheeler
2004a). Vestiture type and density, and leaf texture and degree of toothing may differ between juvenile and adult foliage (Hoogland 1952; Veillon 1990; Toelken 1998). In Hibbertia spp. and Dillenia spp. growing in fire-prone ecosystems, a reversion to juvenile growth is characteristic of fire sprouts.
Vegetative Anatomy. The primary stem anatomy is generally characterized by a uniseriate epidermis, a cortex composed of large, thinwalled parenchyma cells, a continuous cylinder of perivascular fibers and sclereids (absent or scanty in Acrotrema, Didesmandra, Hibbertia spp., and Tetracera spp.), a cylinder of discrete, collateral vascular bundles (or complete vascular cylinder, due to early cambial initiation in Acrotrema, Doliocarpus, Hibbertia, and Tetracera spp.), and a pith containing large, thin-walled parenchyma cells and often also (sometimes mostly) sclerenchymatous idioblasts (Dickison 1970b). Throughout the ground tissue of the stem occur raphid idioblasts and cells with dark-staining contents. Periderm initiation is typically deep-seated, but is sub-epidermal in Dillenia and one Doliocarpus.
Nodes are uniformly trilacunar in Delimoideae, multilacunar (7–27) or trilacunar (Sri Lankan Acrotrema) in Dillenioideae, mostly pentalacunar (with a few spp. 3- or 7-lacunar) in Doliocarpoideae, and uni- or trilacunar in Hibbertioideae; there is one trace per gap. Cotyledonary nodes are unilacunar, two trace, except in Hibbertia, where they are unilacunar, one trace. Dickison (1969) describes petiole anatomy; patterns of petiole vasculature correlate poorly with both nodal type and family-level phylogeny.
Leaves are bifacial. The adaxial epidermis is silicified in Curatella, Davilla spp., Didesmandra, Dillenia spp., and Tetracera spp., and in species of Hibbertia subg. Hemistema, it may be sclerified as well (Dickison 1970a; Rao and Das 1979). Stomata are confined to the abaxial leaf surface and are anomocytic, but in Didesmandra and Tetracera they are paracytic and in Acrotrema they are anisocytic; Hibbertia cuneiformis has cyclocytic stomata. In Hibbertia, stomata may be confined to crypts, or in species with ericoid leaves, confined to two longitudinal grooves, the lamina being recurved almost all the way to the midrib. An adaxial hypodermis is present in Acrotrema costatum, Dillenia spp., and Hibbertia spp. Mesophyll is differentiated into a palisade layer 1–3(4) cells thick and a spongy layer.
Leaves have an unusually diverse array of idioblasts, especially in Hibbertia. Raphid idioblasts occur in all genera except Schumacheria; calcium oxalate crystals also occur as crystal sand and cuboidal or prismatic crystals. Silica bodies are reported from Curatella, Davilla, Doliocarpus, Hibbertia, and Tetracera. Cells described as ‘mucilaginous’ or ‘secretory’, but with uncharacterized contents, occur throughout the family. Tracheoidal or sclerenchymatous idioblasts surround freeending veinlets in many species of Hibbertia (Rao and Das 1979). Trichomes are commonly sclerified, and are occasionally encrusted with silica as well; they are of three basic types.
1. Simple, unicellular trichomes are present in all genera; in Dillenia, they are frequently papillate and highly silicified. Simple trichomes in Acrotrema are sometimes uni- ormulticellular, and raised on a pedestal of enlarged, epidermal cushion cells.
2. Fasciculate trichomes occur in Curatella (Fig. 38), Didesmandra, many species of Hibbertia subg. Hemistema, and Tetracera sect. Tetracera. In species included here in Hibbertia subg. Hemistema (cf.Wheeler 2002), the vestiture consists of (in part) trichomes that are stalked and multiradiate to truly peltate.
3. Short-stalked, glandular trichomes occur on the abaxial leaf surfaces ofDillenia philippinensis and D. reifferscheidia, and on the leaves and sepals of Acrotrema.
For wood anatomy, see especially Dickison (1967a, 1979, 1984), Dickison et al. (1978), and Baretta- Kuipers (1972).Woods typically lack growth rings and are diffuse-porous (excepting a few xeromorphic Hibbertia spp.). Vessels are predominantly exclusively solitary. Vessel elements range in
dimension from c. 70–120 × c. 1,740–2,610 μm in
 |
FIG.38. Dilleniaceae. Curatella americana, various forms of fasciculate hairs on lower leaf surface, x400. (Photograph K. Kubitzki)
|
Schumacheria to c. 150–450 μm× c. 690–1,250 μm in Doliocarpus. Tyloses are infrequently present in the vessel elements of Dillenia spp. only. Intervascular pitting is mostly sparse; pits are circular to elongate,without vestures, and are mostly opposite to transitional (sub-alternate) in arrangement. Perforation plates are exclusively scalariform in Dillenioideae, with 5–160 bars that are completely bordered and occasionally branched. In Hibbertioideae, perforation plates are predominantly scalariform, with 2–42 bars that are frequently branched; simple plates rarely occur, mixed with scalariform plates with few bars, in the woods of a fewHibbertia indigenous to regions with arid climates. In Delimoideae and Doliocarpoideae, perforation plates are mostly simple, with scalariform (1–20 bars, usually less than 10; Doliocarpoideae) or reticulate (Delimoideae) plates occurring only in the smallest vessel elements of a given individual.
Imperforate tracheary (‘fiber-tracheids’) are non-septate, thin- to thick-walled, and very short to very long (625–4,375 μm). These wood fibers have distinctly bordered pits that are common on both radial and tangential walls, and frequently have slit-like, crossed, aperture pairs. Vasicentric tracheids occur in the woods of Tetracera. Axial parenchyma is mostly apotracheal diffuse.
Rays are typically of two distinct sizes. The uniseriate rays are homocellular, consist of upright cells, and are 1–33 cells high. Themultiseriate rays are heterocellular. The ray bodies consist of procumbent to square, infrequently upright, cells; the uniseriate ends are comprised of upright cells and are extended into long wings in Dillenioideae and Hibbertioideae, or in Delimoideae and Doliocarpoideae, may only be one or few cells high.Multiseriate rays vary in width from 1–13 cells in Delimoideae, to 18 cells in Dillenioideae, to 41 cells in Doliocarpoideae, and to 5 cells in Hibbertioideae; in height from 20–152 cells in Delimoideae, 14–200+ cells in Dillenioideae, 32–500+ cells in Doliocarpoideae, and 7–34 cells in Hibbertioideae. Rays of the whole of Dilleniaceae frequently exceed 1mmin height; rays of Doliocarpoideae commonly exceed 10mm in height. The ray parenchyma contains raphid idioblasts in all genera, except possibly Schumacheria; in most New Caledonian Hibbertia, solitary silica bodies commonly occur as inclusions in ray parenchyma cells.
Secondary xylem is without storied structure. In mature, secondary stems of probably all Doliocarpus, lianescent Davilla, Neodillenia, Pinzona, and many neotropical Tetracera, successive, concentric concentric cambia are initiated (apparently rather tardily) that produce interxylary cylinders of secondary phloem, conjunctive parenchyma, raphid idioblasts, islets of thick-walled, rather short sclereids, and in Tetracera, apparently fibers as well.
The early-formed secondary phloem of all genera is composed of sieve tube members, companion cells, scattered parenchyma cells with crystal inclusions, and phloem ray cells (Dickison 1970b). Sclerenchyma is absent. Sieve tube members are usually short (< 300 μm) in all genera of the family. Sieve plates are oblique to occasionally transverse in Delimoideae, Doliocarpoideae, and Hibbertioideae, with typically 2–5 sieve areas, or occasionally the sieve plate is simple. Sieve tube plastids are of the S-type.
Inflorescence Structure. Inflorescences of Dilleniaceae are always determinate. It can be difficult to understand the structure of these inflorescences because of the absence of prophylls (Doliocarpoideae), the production of only a single transverse prophyll (some Tetracera), the recaulescence of many bracts and prophylls (Didesmandra, Dillenia spp., Hibbertia spp., Schumacheria, Tetracera spp.), and the production of serial branches resulting in supernumerary flowers or partial inflorescences (Didesmandra, Dillenia spp., Hibbertia spp.).
Inflorescences of Delimoideae (Tetracera) are characteristically thyrsoids or double-thyrsoids with partial inflorescences consisting of cincinni or dichasia (Kubitzki 1970; sometimes modified and partly monochasial). Inflorescences are terminal and often also axillary, but then only in the axils of the most distal foliage leaves. In cincinni of many neotropical species of sect. Tetracera, one prophyll of each flower is suppressed. In a few species, panicles, botryoids, or dichasia occur. Such inflorescences perhaps represent impoverished thyrsoids. Within Doliocarpoideae, the paniculate inflorescences of Davilla most closely resemble those of Tetracera. They are also both terminal and axillary. In many species, the partial inflorescences are composed of 3-flowered paraclades, each of which is subtended by a bract. The flowers within each paraclade are without prophylls, though the central flower is terminal, and the whole triad appears to develop like a dichasium. Thus, the panicles of many Davilla are basically thyrsoids without prophylls. Subsequent flushes of inflorescences may be produced on the primary inflorescence axis by accessory buds (to a limited extent in Davilla). In large-flowered Davilla and in all other Doliocarpoideae, only a single flower is produced in the axil of each inflorescence bract. In these other Doliocarpoideae, the inflorescences are never terminal (with one possible exception) and are mostly ramiflorous. At each node, subsequent flushes of inflorescences are produced by accessory buds, with nodes producing up to two inflorescence flushes a year for several years. The inflorescences of Curatella and Pinzona are panicles. The inflorescences of Doliocarpus are similar, though less-branched, and perhaps show a reductive trend from an impoverished panicle, to a botryoid, to a monad (Kubitzki 1971). Due to the large number of accessory buds produced by older, leafless nodes of Doliocarpus stems, the inflorescences of this genus are often compound and appear distinctly fasciculate.
In cincinnate inflorescences or partial inflorescences of Dillenioideae and Hibbertioideae, the β-prophyll is displaced onto the branch borne in its axil. Terminal and axillary thyrsoids with cincinnate partial inflorescences are characteristic of Didesmandra and Schumacheria castaneifolia; other Schumacheria spp. have axillary cincinni. Most Dillenia have terminal inflorescences, and in all species they are probably determinate. Cincinni (D. ferruginea and D. triquetra) and thyrsoids (D. albiflos and D. suffruticosa; Corner 1978) occur, though several species have few-flowered cymose inflorescences that need further descriptive work. Solitary, terminal flowers occur in several species with large flowers and are probably impoverished but derived versions of other inflorescence types. The few deciduous species of Dillenia often bloom shortly before leafing out, and produce their inflorescences in axillary or apparently sometimes terminal positions on lateral short shoots (Hoogland 1952). Inflorescences of Acrotrema are either solitary terminal flowers or terminal (and sometimes also axillary), scapose cincinni.
Hibbertioideae are characterized by the prevalence of terminal monads. Indeed, here whole plant architectures may be related to the inflorescence architectures of other subfamilies. Thus, many species of two subgenera of Hibbertia have branching that is prophyllate alone (cymose) for a substantial part of their growth. The case of H. subg. Pachynema has been mentioned (Vegetative Morphology). Species of this subgenus with terete axes show an abrupt, early transition from(1/3)2/5 phyllotaxis to 1/2 phyllotaxis. Prophylls are sometimes the only foliar organs of the adult plant body (Wagner 1906b), which is made up of a compound thyrsoid or paniculate inflorescence. Thus, in H. conspicua, the partial inflorescences (paraclades of first order) are sympodia that terminate in a flower and bear only two prophylls. Hibbertia (Adrastaea) salicifolia, while a wiry scrambler and appearing very different from plants of H. subg. Pachynema, is similarly constructed. Plants undergo an initial phase of extension growth with long shoots producing leaves in a 2/5-spiral, and with flowers confined to (and terminal on) axillary short shoots. These long shoots eventually terminate in a flower. Short shoots of second order may be borne in the axils of leaves of the original short shoots, or axillary branches may be long shoots of a second type, consisting of single-flowered, 2-prophyllate sympodia. In Hibbertia subg. Hemistema and subg. Hibbertia, sympodia often have both an extended zone of vegetative growth with 2/5 phyllotaxis and frequently produce axillary flowers, which obscures the inflorescence-like construction of their plant bodies. Exclusively prophyllate branching is absent as well. Inflorescences of these two subgenera are thus best considered to be terminal and/or
axillary monads (or in aggregation, spiciform or corymbiform polymonads; Toelken 2000), though terminal cincinni and thyrsoids (with cincinnate partial inflorescences resulting from sylleptic, serial branches;Wagner 1906a) also occur in subg.
Hemistema.
Although no comprehensive study of inflorescence structure has been undertaken for Dilleniaceae, the limited investigation undertaken for the preparation of this treatment suggests that inflorescence structure is perhaps the most informative source of structural data enabling the diagnosis of clades above the genus level. Further study is clearly needed.
FloralMorphology. Perianth aestivation is always imbricate, and in 5-merous flowers is quincuncial. The calyx is persistent and slightly to substantially accrescent after anthesis, enclosing the fruit for at least the early stages of its development. However, in Curatella, Pinzona, and some Doliocarpus, the sepals reflex immediately after anthesis and are only slightly if at all accrescent. The corolla is often very showy, and, along with the androecium (and less often the gynoecium), has a prominent role in pollination.Afewspecies of Hibbertia with monosymmetric androecia also have subtly monosymmetric corollas (e.g., H. hyperi coides), but this is variable at the population level. Excepting the above, perianths of Dilleniaceae are polysymmetric.
There is instability in merosity and frequent heteromery of the calyx and corolla in Delimoideae (Tetracera) and Doliocarpoideae, but the mostly uniformly 5-merous perianth in Dillenioideae and Hibbertioideae is probably derived. In Delimoideae, the calyx is typically 4–5-merous, with the number of petals often equal to the number of sepals. Several species of Tetracera, however, possess flowers with an unequal calyx of two small outer sepals and three large inner sepals, and have only three petals alternating with the inner sepals. At least five species of Tetracera have a calyx of 7–15 helically arranged sepals; their corolla merosity (3–5) is apparently related to the number of orthostichies in the sepals. Doliocarpoideae are the most variable subfamily with respect to perianth merosity; heteromery is frequent.Within Dillenioideae and Hibbertioideae, deviations from pentamery are almost certainly derived. Calyces of 7–18 sepals occur in at least six species of Dillenia, while a corolla is absent in a few other species. While the calyx of Hibbertia is uniformly 5- merous, in five New Caledonian species (formerly sect. Trisema) the corolla is often only 3-merous.
The androecium has mostly 10–300 stamens, although a few Hibbertia have as few as 1–5, some Tetracera have 500, and Dillenia ovalifolia has up to 900 stamens. Polymerous androecia may be plesiomorphic for Dilleniaceae, and androecia with fewer than 11 stamens or greater than 200 stamens appear to be derived. Androecia of Delimoideae and Doliocarpoideae are uniformly polysymmetric. Constitutional androecial monosymmetry occurs in Dillenioideae (Didesmandra and Schumacheria) and Hibbertioideae (most species of Hibbertia subg. Hemistema), and is independently derived within each of these subfamilies. Except in a fewHibbertia species, thismonosymmetry is conspicuous, manifested by the formationof only a single group (or 2 groups in Didesmandra) of fertile stamens between the gynoeciumand the petal that is presented inthemedian plane of the flower.Thus, the group(s) of fertile stamens is (are) presented either abaxially or (less commonly) adaxially.
The androecium is always without staminodes in Delimoideae and Doliocarpoideae, whereas in many species of Dillenioideae (many Dillenia) and Hibbertioideae (species of all subgenera of Hibbertia except Adrastaea), there are also staminodes. Staminodes are±filamentous in form(infrequentlywith
infrequentlywith a vestigial anther) and are as long as to, commonly, much shorter than the fertile stamens. In polysymmetric androecia, staminodes are most commonly external to the stamens, though in some species of Hibbertia subg. Pachynema and Dillenia fagifolia they are internal. In species of Hibbertia subg. Hemistema with monosymmetric androecia, staminodes may be external to the group of fertile stamens (e.g., sect. Hemistema), lateral to them (numerous spp.), or may partly or entirely encircle both androecium and gynoecium as a ± uninterrupted ring (e.g., H. hypericoides, H. spicata).
The stamens are characteristically entirely free. In polysymmetric androecia, the stamens are evenly distributed around the gynoecium, either arranged without apparent order or distinctly antesepalous. In Hibbertia (subg. Adrastaea) salicifolia alone, the androeciumis obdiplostemonous (Tucker and Bernhardt 2000; cf. Baillon 1865). Androecia sectoralized into stamen fascicles the members ofwhichare freeoccur in Acrotrema spp., and in both polysymmetric and monosymmetricflowered species of Hibbertia. More commonly, there is basal to nearly complete connation of the filaments in each fascicle, as in Didesmandra, Hibbertia (many species of subg. Hemistema and most species of subg. Hibbertia sect. Candollea), and Schumacheria. A single stamen fascicle occurs in many species of Hibbertia subg. Hemistema and in Schumacheria; there are two fascicles in Didesmandra and three or five fascicles in Hibbertia sect. Candollea. In Hibbertia salicifolia and some species of both subg. Hibbertia and subg. Pachynema, the filaments of the outer whorl of stamens are basally connate, forming a short tube.
Stamens of all Dilleniaceae have clearly differentiated anthers and filaments. Filaments are typically terete and vary frombeing c. 10×shorter than the anthers, as in some Dillenia, to c. 10× longer than the anthers, as in a few species of Tetracera and Davilla. The filaments of Tetracera gradually dilate in their distal 1/4, grading into the connective. In Hibbertia subg. Pachynema, filaments may be dorsiventrally flattened (H. conspicua), or frequently have an expanded, bulbous base that distally abruptly tapers into a narrow, terete cylinder. Anthers are always basifixed, dithecal and tetrasporangiate. They commonly dehisce via latrorse or (infrequently) introrse longitudinal slits, but inDillenioideae andHibbertioideae anther dehiscence is frequently via apical pores or clefts. The apically porose anthers in many Dillenia are extrorse (Endress 1997; but not always, cf. Hoogland 1952).
Anthers range from slightly wider than long and c. 1mm in length in Tetracera spp. to the narrow, linear anthers of some Dillenia that are over 20mm in length. In Tetracera and many Doliocarpoideae (especially Davilla), the anther connective is laterally broadened, separating the thecae especially toward the base. Other Doliocarpoideae (Curatella; some spp. of Doliocarpus sect. Doliocarpus, especially D. grandiflorus and D. magnificus; Neodillenia) have anthers with parallel thecae and a rather narrow connective, and approach in form those of Hibbertioideae and Dillenioideae. These latter nearly all have parallel thecae (slightly basally divergent in a few Hibbertia subg. Pachynema) and narrowconnectives. In Didesmandra,Dillenia spp., Hibbertia spp, Tetracera spp., and Schumacheria, the connective apically protrudes as a short mucro. In many Dillenia, there is an internal group of stamens (which are reflexed in bud) with anthers that are much longer and sometimes differently colored than those of the external stamens. In several New Caledonian Hibbertia (Veillon 1990), the internal stamens have large, often red or lavender, falcate anthers that are distinctly larger than the ± straight, often yellow anthers of the outer stamens, while in Didesmandra each stamen fascicle consists of one large stamen with a long, apically uncinate anther and four smaller stamens with comparatively short, straight or slightly curved anthers.
The gynoecium is characteristically apocarpous and hypogynous, and excepting Dillenia, nearly always consists of 1–5 carpels; the gynoecia of Dillenia (with (4)5–15(–20) carpels), Hibbertia grossulariifolia (with (5–)10(–15) carpels), and some populations of H. scandens (with up to 7–8 carpels) are clearly derived. Carpels are arranged in a single whorl, except in Hibbertia grossulariifolia, where two distinct whorls are present (Tucker and Bernhardt 2000). Carpels are most frequently entirely free or shortly basally connate in the ascidiate region of each carpel; there is no compitum. However, the gynoecia of Acrotrema, Curatella, many (perhaps all) Dillenia, and Pinzona are hemisyncarpous – a syncarpous zone occurs where pollen tubes may cross between carpels. Stylodia are always free, and are erect, or in some Dillenia and Hibbertia, are laterally exserted. In Dillenia and also apparently Hibbertia baudouinii (Wilson 1965), the carpels are basally adnate to the floral receptacle, which is conical in the region of the gynoecium and often apically protrudes from the center of the whorl of carpels. Endress (1997) interprets this as a byproduct of a secondary increase in carpel number. The flowers of Hibbertia grossulariifolia, H. lasiopus, H. miniata, H. montana, and H. quadricolor (all immediately related; J.W. Horn, unpubl. data) are perigynous.
Thecarpelshaveaveryshortascidiatezoneand a large, well-developed symplicate zone. The ventral suture of each carpel, which is always conspicuous and extends to the tip of (or sometimes just below) the stylodium, is sometimes not fully (morphologically) sealed (Dickison 1968). The stigma is punctiform, minute, and not differentiated in shape fromthe stylodium (or rarely minutely capitate) in Dillenioideae and Hibbertioideae. Stigmas of Doliocarpoideae are peltate, the even margin being interrupted only by the ventral suture of the carpel. In Delimoideae, stigmas are mostly punctiform or otherwise small and indistinct, though in a few neotropical Tetracera they are peltate, but with irregular, somewhat jagged margins.
Placentation is submarginal with ovules in two (sometimes four in Dillenia, or to six in Tetracera; Dickison 1969) vertical rows when there are four or more ovules in each carpel. Species with only 1–2 ovules per carpel have basal placentation. Ovule number per carpel ranges from 1, as in Didesmandra and Schumacheria, to c. 80 in some Dillenia; Doliocarpoideae characteristically have two ovules per carpel. Ovules at anthesis are anatropous to campylotropous. When ovules are numerous, they are pleurotropous (borne sideways) and more or less syntropous (curved with the carpel margin). When there are 1–2 ovules per carpel, they are always erect and may be uniformly apotropous (abaxially curved), or, in most Doliocarpoideae, with one ovule apotropous and the other epitropous (adaxially curved).
Flowers are oriented so that theirmedian petal is abaxial. Carpels are opposite the petals when isomerous with the corolla, (commonly) median when two, and in 3-carpellate species of Schumacheria, the odd carpel is abaxial-median. The single carpel of many species of Davilla and Doliocarpus is variably obliquely oriented. While the bicarpellate gynoecia of Hibbertia subg. Adrastaea and subg. Pachynema are clearly median, species of subg. Hemistema with two carpels present their flowers such that the carpels are transverse, as does Didesmandra. In fact, monosymmetric flowers of Dilleniaceae are presented 90◦ from their true morphological orientation, the plane of symmetry of the flower being transverse (cf. Eichler 1878); this is most clearly seen in species with cincinnate inflorescences. The obliquely oriented, bicarpellate gynoecia of Didesmandra may be caused by the loss of the (as presented) median carpel of an ancestrally 3-carpellate flower, as in Schumacheria (Stebbins and Hoogland 1976) – indeed, Schumacheria and Didesmandra are sister taxa. However, the oblique orientation of the carpels in species of Hibbertia subg. Hemistema is independently derived there and is expressed late in development (Tucker and Bernhardt 2000).
Floral Anatomy and Development. Sepals are (fundamentally) vascularized by three traces, with the ateral sepal traces commonly commissural, or there is a single trace from the stele that branches into three traces below the sepal base, or remains unbranched (some Hibbertia; Sastri 1958). Calyx vascularization in Delimoideae and Doliocarpoideae is frequently irregular (Wilson 1973). The petals are each vascularized by a single trace that is either fused with a commissural lateral sepal trace or is discrete. Androecial vasculature is typically discrete from that of the perianth, with the traces to the stamens almost always basally fused into (1–)3–15(–25) trunk bundles (Wilson 1937, 1965, 1973). The stamen trunk bundles typically branch 2–3 times, with the individual stamens or staminodes each receiving a single trace. Rarely, as in Hibbertia subg. Pachynema, each stamen receives its trace directly from the stele. Gynoecial vasculature is separate from that of the androecium, with each carpel characteristically vascularized by 3 traces – a single dorsal trace and 2 ventral traces. In Curatella and Doliocarpus spp., a pair of accessory ventral traces is present that supply the ovules. Ovules are otherwise vascularized by branches from the carpel ventrals, or in Didesmandra and Schumacheria, by vascular tissue remaining in the floral receptacle after the departure of the carpellary traces (Dickison 1968). In Dillenia spp. and Hibbertia spp., residual stelar tissue remains in the floral receptacle above or internal to the point where the carpel ventrals depart.
Floral development is known mainly in Hibbertia and Dillenia (Endress 1997; Tucker and Bernhardt 2000; and references therein). The sepals are always spirally initiated (2/5 when 5), and in Dillenia, long plastochrons occur between the initiation of each sepal. The initiation of the corolla follows, with the petal primordia arising simultaneously or nearly so (many Hibbertia spp.), or initiated in the samephyllotactic spiral as the calyx (Dillenia; H. salicifolia). In species of Hibbertia subg.
Hemistema with monosymmetric androecia, the three petals on the same side of the flower as the fertile stamens are initiatedbefore the twoontheother side of the flower. The innermost stamen primordia are typically next to arise, and are commonly antesepalous, though in species of Hibbertia subg. Hemistema with monosymmetric androecia, they are±opposite the median petal. These first stamen primordia appear on either a ring meristem (Dillenia, H. salicifolia, H. scandens) or on ± discrete, common primordia that may be spirally initiated (other polysymmetric-flowered Hibbertia spp.; cf. H. perfoliata, Tucker and Bernhardt 2000). Additional stamen primordia are initiated centrifugally, with concomitant growth of the floral receptacle, increasing its diameter, in species with particularly large numbers of stamens. In Tetracera nordtiana, the developing androecium also has a minor centripetal zone, with a few stamen primordia arising in sequence towardthe centerof theflower (Endress 1997).Thecarpelprimordia initiatesimultaneously (or in H. grossulariifolia, the internal carpel primordia after the external), most commonly when stamen primordia are being centrifugally initiated. In Tetracera, the carpel primordia arise prior to the initiation of individual stamen primordia (Corner 1946; Endress 1997). In monosymmetric-flowered species of Hibbertia subg. Hemistema, the first initiated stamen primordia arise as a±terminal ridge on the floral apex. In fact, subsequent to the initiation of the corolla, the floral apex functionally splits in the transverse plane of the flower (at least as the flowers are presented).The side adjacent to themedian petal gives rise to the primordia of the fertile androecium (sometimes also staminodes) and the other half gives rise to the carpel primordia (rarely also staminodes).
Later in floral development, the stamens are either straight, contorted (most of the family), or become
partly reflexed (several Dillenia species with a heterantherous fertile androecium, see above; Corner 1946; Hoogland 1952; Endress 1997) or wholly reflexed in bud (Doliocarpus sect. Doliocarpus, Kubitzki 1971).
Embryology. Embryological investigations have focused on only three genera and few species (Paetow 1931; Swamy and Periasamy 1955; Rao 1957; Sastri 1958; Lakshmanan and Lakshmanan 1984; Imaichi and Kato 1996); Delimoideae and Doliocarpoideae are embryologically almost unknown. The immature microsporangium wall is 4–5 cell layers thick, and middle layers become crushed during development. The mature microsporangium wall is unusual in that the epidermal layer is always most prominent, the cells typically being tanniniferous or, at least, with dark-staining contents. The endothecium is often poorly developed; fibrous thickenings are absent in Dillenia spp., inconsistently developed in Acrotrema arnottianum and several Hibbertia spp., but apparently consistently present in H. stricta. The tapetum is probably of the secretory type, and microsporogenesis is simultaneous. Pollen grains are shed as monads at the 2-celled stage.
Anthetic ovules are anatropous to campylotropous, bitegmic, crassinucellate, and have a zigzag or, less often, exostomal micropyle. The outer integument is hood-shaped, and is 2–3 cells thick at anthesis. The inner integument is 3–4 cells thick at anthesis. Both integuments are thickened in the region of the micropyle. The raphe is often short, especially in Doliocarpoideae. Nucellar cells in the region of the chalaza retain their meristematic potential and continue to divide during a period from megagametophyte development to a point well after fertilization; this extended nucellar growth is probably primarily responsible for the eventual conspicuous curving of the ovule body. After fertilization, cells in the region of the raphe also continue to divide (becoming multinucleate in A. arnottianum, Swamy and Periasamy 1955), and this region grows inward against the nucellus and developing endosperm, which perhaps also helps effect the eventual curvature of the ovule body. Hence, post-anthetic ovules become truly amphitropous in all species examined. Spiral thickenings appear in cells of the outer epidermis of the inner integument at the time of fertilization, or shortly after.
The mature megagametophyte is 7-celled and 8-nucleate, and its development is of the Polygonum type. The synergids are obpyriform in shape, or in Hibbertia, are hooked. The polar nuclei fuse before fertilization, and the three antipodal cells are ephemeral in Hibbertia and Dillenia. The primary endosperm nucleus divides before the zygote; endosperm formation is Nuclear. Embryogeny probably follows the Onagrad type. A zygotic mantle, formed by the swelling and structural modification of the zygotic membrane, is reported from Acrotrema, Dillenia, and Hibbertia, and persists during early embryogeny, sometimes projecting with irregular, finger-like protuberances among the adjacent endosperm cells (Swamy and Periasamy 1955; Sastri 1958; Ioffe and Zhukova 1974). This unusual elaboration of the zygotic membrane has been reported from all three genera embryologically examined, and is apparently restricted to the family.
Pollen Morphology. Pollen grains are mostly spheroidal in shape, though they may vary from subprolate to suboblate. Grains vary in size from 15–34×16–32 μm,with the largest grains observed in Hibbertia stricta (34 × 32 μm), and the smallest in Schumacheria castaneifolia (15 × 16 μm; Dickison 1967b). Exine structure is tectate-perforate to semitectate, with finely punctate or foveolate to coarsely reticulate sculpturing. Exines of Davilla and Doliocarpus are the most coarsely reticulate, andvery finely punctate exine sculpturingoccurs in pollen of all subfamilies recognized here (Dickison et al. 1982). The tectum is typically well-developed, accounting for up to one-third of the total thickness of the exine, and lies above a layer of prominent, infratectal bacula. The foot layer and endexine region of the nexine (nexine 1 and 2) are often poorly differentiated from one another. The sexine is as thick as to c. 4× as thick as the nexine; the nexine is, however, considerably thickened in the region of the apertures in all taxa (Dickison et al. 1982). Viable pollen grains are triaperturate in all genera, except Didesmandra and Schumacheria in which the grains are mostly tetracolpate. Apertures of Delimoideae and Doliocarpoideae are compound(colporate),with mostly circular endoapertures that are sometimes poorly differentiated from the colpal membrane. Apertures in Dillenioideae andHibbertioideae are simply colpate; endoapertures have evidently been lost in these two subfamilies. The length of the colpi varies from very short in some Dillenioideae to very long in some Doliocarpoideae. In the neotropical species of Tetracera, which are structurally androdioecious, the pollen is strikingly dimorphic (Kubitzki 1970). Plants bearing only staminate flowers produce conventional tricolporate pollen, whereas plants with structurally perfect flowers produce pollen that is indistinctly 5–8-pantoporate tonearly inaperturate (‘cryptoporate’)andalwaysempty, obviously forming a deceptive attractant for the visitors of the functionally female flowers.
Pollination. Early investigations (Keighery 1975; Gottsberger 1977) suggested beetles were the primary pollinators of flowers of individual species of Hibbertia, as well as those of Dilleniaceae as a whole. However, more recent work indicates that their flowers are principally bee pollinated. As the flowers are apparently uniformly without nectar, pollen serves as the sole reward. In Dillenia spp. and Hibbertia spp. with (at least functionally) apically porose anthers, bees actively remove pollen from the anthers via thoracic vibrations (Bernhardt 1984, 1986; Keighery 1991; Endress 1997; Tucker and Bernhardt 2000). In Hibbertia, bees sometimes also use their forelegs to scrape pollen from the anther pores (Bernhardt 1986). Tucker and Bernhardt (2000) outline four pollinator syndromes in Hibbertia that are related to the diverse floral architectures in this genus. Thus, although different species of Hibbertia (H. fasciculata and H. stricta)may be pollinated by bees of the same genus, pollen is deposited in different places on the bees’ bodies from where it is picked up by stigmas that intercept the bees on the appropriate parts of their bodies (Bernhardt 1996). Hence, variation in floral architecture among sympatric Hibbertia species may function as an interspecific isolating mechanism (Bernhardt 1996). While bees of the families Colletidae and Halictidae are apparently the primary pollinators of Hibbertia spp., pollen-eating flies of the family Syrphidae may be secondary pollinators in some species (Bernhardt 1996; Tucker and Bernhardt 2000).
Endress (1997) reports revolver flowers and roundabout flowers, correlated with two distinct pollination modes (both ofwhich involve buzz pollination by Xylocopa bees) in Dillenia spp. (mostly observed in cultivation). Observations of the pollination biology of Dillenia spp. in native habitats also indicate visitation or pollination by bees (Apis, Ceratina, Melipona, and especially Xylocopa; summarized by Endress 1997;Momose et al. 1998). The scarce reports on pollination biology of other Dilleniaceae also indicate visitation by bees; Tetracera akara is visited by Apis koschevnikovii (Momose et al. 1998), and Davilla nitida is visited by Trigona sp. (Croat 1978).
Fruit and Seed. Fruits are most commonly follicles or aggregates of follicles. In Dillenia spp. with dehiscent fruits and Acrotrema, the fruitsmay technically be considered capsules on account of the hemisyncarpous gynoecia of these genera. They are, however, probably best described as an aggregate of basally coherent follicles dehiscing along the ventral suture in Dillenia, or irregularly in Acrotrema. In several species of Dillenia, the pericarps remain drywhile the accrescent sepals,which become fleshy, form a tough, persistent envelope around the fruit (Fig. 48E, F); these are called pseudocarps (Hoogland 1952). In all species of Davilla, the function of the fruit wall, which remains very thin, is taken over by the two innermost sepals, which are accrescent and enclose the fruit completely like a pair of clam shells. The two accrescent sepals become distinctly cartilaginous and often turn orange at maturity; the whole accessory fruit thus resembling a globose capsule. Baccate or
leathery, capsular fruits occur in Curatella and Pinzona. In these genera, the dorsal part of each carpel growsmore than the ventral part, so in fruits nearing maturity, the stylodia are crossed. The fruits of Doliocarpus are also fleshy, and are either fleshy follicles that are dehiscent (sometimes irregularly) along both the dorsal and ventral portions of the carpel, or indehiscent and berry-like; the pericarps typically ripen red. In many species of Acrotrema, Dillenia, Hibbertia, andTetracera, several seeds per carpel reach maturity, whereas in most other genera, carpels are 1–2-seeded.
Funicular arils are present on the seeds of all genera. However, the aril is vestigial in many Dillenia spp.with indehiscent fruits and is represented onlyby a slight annular thickeningonthe funiculus. The cells of the aril of Acrotrema arnottianum are reported to be coenocytic (Swamy and Periasamy 1955). Arils are red,white, or (rarely) orange, fleshy, and typically oily or waxy, or in many Hibbertia spp., are scarious, beige, and waxy. The margin of the aril varies from being deeply incised with long, thin laciniae (most Tetracera) to shallowly incised (many Dillenia, Hibbertia cuneiformis, and H. scandens), or is subentire to undivided in all other genera. The aril may be asymmetrical.
Seeds are spheroidal to reniform and are characteristically of a lustrous black or dark brown color, owing to the presence of pigments and tannins in the exotesta. The exotesta of many New Caledonian Hibbertia is red. The testa is 2(–4) cell layers thick and is not multiplicative. The exotesta is always unlignified and may be slightly pulpy or, in at least some Davilla and Dillenia spp., it dries up. In some Acrotrema and Dillenia, the exotesta is provided with slender, unicellular trichomes that may be mucilaginous. The principal mechanical layer of the seed coat is the endotesta, whose cells are heavily lignified and strongly pitted, or in some Hibbertia are heavily cutinized (Corner 1976; Schatral 1995). In Tetracera, the endotesta is poorly differentiated fromother layers of the testa overmuch of the seed. Many of the cell layers of the inner integument of the ovule become crushed during seed development; hence, the tegmen is typically only 1–2(–4) layers thick. The exotegmen is distinctive as its cells have spiral or annuliformthickenings in all genera examined (i.e., a tracheidal exotegmen; Corner 1976; Vyshenskaya and Oganezova 1991). The chalaza is massive; its cells are often lignified or cutinized. The raphe is short. Endosperm is nuclear, abundant, fleshy and oily, or in Hibbertia, also with starch granules. The embryo is straight, oftenveryminute,andnot fully differentiatedwhen the seed is mature. Germination is phanerocotylar (Ozenda 1949).
Dispersal. Seed dispersal is probably mostly by animals. Birds probably disperse the arillate seeds produced by species of Delimoideae, Doliocarpoideae, and Dillenioideae with dehiscent fruits. The ornithochorous syndrome is well-displayed by many species of Dillenia and Tetracera, in which a red aril contrasts with the shining, black testa, and a follicle colored in a different red. In Curatella and Doliocarpus, the combination of the red color of the pericarp, white aril, and black testa provides a visual display that is also probably attractive to birds. Only few species of Hibbertia have red arils and are ornithochorous (H. scandens and H. cuneiformis), butmany others are myrmecochorous (Berg 1975; Schatral et al. 1994). The large, indehiscent fruits of several Dillenia, which have accrescent sepals and lack arils, are eaten by mammals; D. indica, specifically, is often eaten and dispersed by elephants, but also transported by water.
Reproductive Biology. Flowers of Dilleniaceae (despite their sometimes very large size) have a short period of anthesis, only one to few days (Endress 1997; Tucker and Bernhardt 2000). Hybridization within Hibbertia, and perhaps throughout the whole family, is apparently extremely rare (Toelken 1998; Tucker and Bernhardt 2000). Hibbertia hypericoides is protogynous, and this and other species of the genus are self-fertile, although autogamy does not seem to occur (Keighery 1975). The functional dioecy present in the neotropical species of Tetracera (see Pollen Morphology) enforces cross-pollination. The flowers of Hibbertia hirsuta typically possess only a single stamen, and are probably cleistogamous.
Phytochemistry. Polyphenols are prominent and diversified and include hydrolyzable tannins and ellagitannins, while alkaloids seem to be rare, if present at all (Hegnauer 1966). Betulinic acid, mostly accumulated in the bark, appears to be a regular feature of the family (Pavanasasivam and Sultanbawa 1974). The diversity of flavonoids is high, and besides the common aglycones and glycosides, some unusual O-methylated compounds plus flavonol sulphates have been found, the latter two, in a close association (Kubitzki 1968; Gurni and Kubitzki 1981).
Relationships Within the Family. The principal characters used for earlier subdivisions of Dilleniaceae include anther structure (linear vs. globose), leaf architecture (lateral nerves welldeveloped and often parallel vs. small, sometimes 1-nerved leaves), anther dehiscence (longitudinal slits vs. apical pores), and degree of carpel fusion (carpels entirely free vs. ± synovarious). Their application has led to three slightly different subdivisions: (1) that of de Candolle (1824), who recognized two tribes, Delimeae and Dillenieae, which Hoogland (1952) informally elevated to the rank of subfamilies; (2) that of Gilg and Werdermann (1925), into the tribes Tetracereae, Hibbertieae, Acrotremeae, and Dillenieae; (3) the division of Hutchinson (1964),whomodified Gilg andWerdermann’s classification by merging Acrotremeae into Dillenieae. Molecular phylogenetic data are highly informative as to the infrafamilial relationships of Dilleniaceae, and largely corroborate the classifications based on structural data mentioned above. Following Horn (2005; Fig. 39), Dilleniaceae are best divided into the subfamilies Delimoideae, Doliocarpoideae, Hibbertioideae, and Dillenioideae. Delimoideae, here taken to include only the genus Tetracera, are sister to the rest of the family. Doliocarpoideae, containing the neotropical endemic genera Curatella, Davilla, Doliocarpus, Neodillenia, and Pinzona, are sister to a clade containing the Old World genera. The Old World clade contains the sister cladesHibbertioideae, here taken to include only the broadly circumscribed Hibbertia (Adrastaea and Pachynema included), and Dillenioideae, containing Acrotrema, Didesmandra, Dillenia, and Schumacheria.
 |
| Fig.39. Dilleniaceae. Summary tree of relationships in Dilleniaceae. The branches shown here have bootstrap support of ≥ 99% and Bayesian posterior prrobability values of 100%, except those indicated with a dashed line, where bootsrtrap values sre <70% and Bayesian posterior probability values are <95% (from Horn 2005). |
Morphological circumscriptions of major clades of Dilleniaceae are indicated in the taxonomic
section. Characters supporting hypotheses of relationship for both Didesmandra and Neodillenia,
which were not available to include in the phylogenetic study, are also indicated in
this section. The clade containing all genera of Dilleniaceae, exclusive of Tetracera, is strongly
supported by molecular data (Horn 2005), but as yet no structural characters have been identified
to diagnose this group.
Affinities.
In previous systems of angiosperm classification (Dahlgren 1983; Cronquist 1981, 1988; Takhtajan 1997; Thorne 2000), Dilleniaceae were accorded a position of great evolutionary importance as being, on one hand, directly liked with magnoliid dicots, and evenmore significantly on the other, as being the progenitor of an entire radiation of dicots that were, in part, characterized by their centrifugal stamen initiation – the Dilleniidae
Dilleniidae.
Part of this confused notion is attributable to a problem that still stands: Dilleniaceae are one of the most isolated lineages of eudicots of which the relationship to other major eudicot lineages cannot (currently) be robustly supported by any source of character information. Dilleniaceae are currently most conservatively treated as an unresolved, early eudicot branch, and are accordingly contained within the monofamilial Dilleniales (APG II 2003).
There is, however, a growing consensus of molecular data suggesting Dilleniaceae are sister to Caryophyllales (Soltis et al. 2003). Dilleniaceae and many Caryophyllales share deep-seated phellogen initiation, successive cambia, persistent calyces, and campylotropous ovules. Within Caryophyllales, Dilleniaceae are perhaps most similar to Rhabdodendraceae, probably sister to all other Caryophyllales (Cuénoud et al. 2002). Both families share the following character states of limited distribution among eudicots: leaf mesophyll containing silica bodies, a broad petiole with at least a somewhat sheathing insertion, anthers with a persistent, tanniniferous epidermis, and a seed coat with a tanniniferous outer epidermis and a layer or layers of cells developing spiral thickenings (for Rhabdodendraceae: Prance 2003).
Distribution. Tetracera, the sole genus of subfamily Delimoideae, is the only genus with a pantropical distribution, as well as the only genus of the family present on continental Africa. Doliocarpoideae, containing 4–5 genera, are endemic to the neotropics, with a center of diversity in Brazil. Hibbertioideae are mostly endemic to Australia, and are the largest subfamily, perhaps containing more species than the rest of the family as a whole. Their distribution in Madagascar (1 sp.), New Caledonia (24 spp.), Fiji (1 sp.), and New Guinea (2 spp.) is the result of long distance dispersal, rather than vicariance (J.W. Horn, unpubl. data). Hibbertioideae have a significant temperate distribution in southern Australia, and the Southwest Botanical Province of Western Australia is the region of greatest species richness for both the subfamily and, perhaps, for the entire family as well. Dillenioideae have a distribution from Madagascar and the Seychelles to Sri Lanka and southern and eastern India, to Southeast Asia and the Pacific (northern Australia and Fiji). The distribution of Dillenia is equivalent to that of the subfamily. Schumacheria is endemic to Sri Lanka. Didesmandra is known from only a few populations in Sarawak, Borneo. Acrotrema contains seven species endemic to Sri Lanka, one species in theWestern Ghats of India, and another ranging from southern Thailand and Myanmar to northern Sumatra. The biogeographic history of Dillenioideae is probably congruent with the out-of-India hypothesis (Ashton and Gunatilleke
1987; Conti et al. 2002).
Key to the Genera
1. Fertileandroeciumexclusivelyonone sideof theflower 2 – Fertile androecium ± evenly distributed around the carpels 4
2. Stamens 10, grouped into 2 fascicles, each of 5 connate members; (fertile) androeciumconspicuously heterantherous 8. Didesmandra – Stamens 1–c. 70, with fertile stamens clustered into a single fascicle; anthers ± isomorphic, stamen filaments free or fused 3
3. Carpels (2)3; staminodes 0; stamen filaments connate; anthers with fine, simple hairs; flowers in secund, cinncinate inflorescences or partial inflorescences 7. Schumacheria – Carpels 2; staminodes present or not; stamen filaments free or connate; anthers glabrous; inflorescences typically of a single flower, or the few species with flowers organized into secund, cinncinate inflorescences always bearing flowers with staminodes 6. Hibbertia (p.p.)
4. Stigma peltate, the margin even and annuliform. Inflorescences terminal, axillary, or (frequently) ramiflorous, prophylls absent or (very rarely) minute. Aril always well-developed, entire or shortly and irregularly lacerate distally, typically fleshy. Lianas and scandent shrubs, less commonly shrubs or tortuous trees 5 – Stigma punctiform, rarelyminutely capitate, or if irregularly peltate (a fewTetracera), then the plantswith terminal, prophyllate inflorescences and laciniate arils. Inflorescences commonly terminal; 1–2 prophylls/flower. Arils well-developed or vestigial, fleshy or scarious, entire to deeply fimbriate or laciniate. Large trees to shrubs or lianas, rarely rhizomatous herbs 9
5. Sepals uniformly 5, the inner 2 opposite one another, deeply concave, and distinctly larger than the outer 3 ± flattened sepals; the two inner sepals accrescent, enclosing the fruit completely; carpels 1–2(3); inflorescences terminal and axillary, a panicle, commonlywith 3 flowers per bract 2. Davilla – Sepals 2–7 (frequently 5), ± equivalent in size and shape, or if unequal, then the inner 2 sepals not as above, either all sepals are accrescent and ± surround the fruit, but never completely, or they reflex, becoming at most slightly accrescent; inflorescences axillary and/or (commonly) ramiflorous (very rarely terminal), a panicle (always with 1 flower per bract), fascicle of panicles or botryoids, or rarely flowers single; carpels 1–2(–5), gynoecium apocarpous or hemisyncarpous 6
6. Carpels 2, hemisyncarpous 7 – Carpels 1, rarely 2–5, the carpels free or atmost shortly synovarious 8
7. Small tomedium-sized, tortuous trees; hairs both simple and fasciculate; carpelswith long, persistent, hispid hairs; aril white 3. Curatella – High-climbing lianas; hairs simple only (plants often
mostly glabrous); carpels glabrous; aril orange 4. Pinzona
8. Aril white; carpel 1(2); inflorescences exclusively axillary and ramiflorous 5. Doliocarpus – Aril red; carpels 1–5, if carpel 1, then the inflorescence terminal 5a. Neodillenia
9. Anthers with a conspicuously expanded connective separating the thecae; thecae basally outwardly divergent; stamen filaments increasing in thickness in distal 1/4, grading into the connective 1. Tetracera – Anthers with a narrow connective, the thecae often immediately next to one another and always parallel; stamen filaments of even thickness throughout their lengths, or rarely with an expanded and bulbous base and then abruptly narrowing 10
10. Rhizomatous herbs with a single rosette of leaves at ground level, or uncommonly with a short, aerial nonphotosynthetic stem that is densely pubescent and terminated by a rosette of leaves 9. Acrotrema – Large trees to small shrubs, or rarely subshrubs or lianas; if plants are subshrubs, then the above ground axes are green and photosynthetic, ephedroid or conspicuously flattened, and appearing leafless 11
11. Portion of floral receptacle bearing the carpels conical; carpels (4)5–15(–20), gynoeciumhemisyncarpous; petiole bases with 7 or more vascular bundles; leaves moderate in size to typically large and broad (never long and strap shaped), with ± straight, parallel secondaries andfrequently regularlypercurrent tertiaries; areoles typically well-developed 10. Dillenia – Portion of the floral receptacle bearing the carpels flat, or if conical, then the leaves long and strap-shaped; carpels 1–5(–15), entirely free, or at most shortly synovarious; petiole bases (or lamina bases if blade is sessile) with 1 or 3 vascular bundles; leaves of moderate to very small size, sometimes ericoid, uncommonly (mostly) only scale-like and the plant appearing leafless; tertiary venation rarely percurrent and never regularly so; areoles typically poorly developed 6. Hibbertia (p.p.)
Genera of Dilleniaceae
Plesiomorphic states:
Vessel elements with mostly simple perforation plates.Multiseriate wood rays with uniseriate ends both fewer and greater than 4 cells in height (Kribs Type I and IIA). Leaves without completely amplexicaul petiolar wings, but sometimes the leaf insertion with a broad, slightly sheathing petiolar flange.Venationcraspedromous or semicraspedromous; the secondaries ± straight and parallel, the tertiaries rigidly percurrent and scalariform, areoles well-developed. Vestiture exclusively of simple, unicellular trichomes. Inflorescences terminal, determinate, not branching from the previous flush via proleptic proliferation buds. Androecium polysymmetric. Functional pollen grains tricolporate. Gynoecium apocarpous, or only very shortly synovarious by the ascidiate region of each carpel and without a compitum. Stigmas small and punctiform, not differentiated in shape from the stylodia. Fruit a follicle or aggregate of follicles.
I. Subfam. Delimoideae Burnett (1835).
1. Tetracera L.
Fig. 40
Tetracera L., Sp. Pl. 1:533 (1753); Hoogland, Reinwardtia 2:185–224 (1953), rev. Australasian spp.; Kubitzki, Mitt. Bot. Staatssamml. München 8:1–98 (1970), rev.; Hoogland, Fl. Thailand 2, 2:95–108 (1972); Aymard in Fl. Venez. Guayana 4:671–685 (1998).
Lianas or scandent shrubs, rarely lignotuberous subshrubs; mature, secondary stems often with successive cambia; small vessel elements with reticulate plates; nodes uniformly trilacunar, 3-trace; stomata paracytic; trichomes often fasciculate. Plants synoecious or functionally dioecious. Inflorescences mostly both terminal and in the axils of the upper foliage leaves, very rarely exclusively axillary, with (1–)3–150(–200+) flowers, a thyrsoid or double thyrsoid with either (modified) dichasial or cincinnate partial inflorescences, rarely a dichasium or impoverished panicle; prophylls 1–2/flower; sepals (3)4–5(–15), free, unequal to equal; petals 3–5; stamens 50–200(–500), free; anthers with a short, mucronate appendage or not; the connective emarginate or not; carpels 1–5(–8), glabrous or pubescent; stigmas infrequently irregularly peltate with jagged margins; ovules 2–20, pleurotropous and syntropous, in 2(4, 6) submarginal rows. Follicles pyriform. Seeds 1–6 per follicle, arillate; aril fleshy, red (drying beige), ± evenly fimbriate or laciniate in its distal (1/8–)1/3 to (more commonly) nearly its full length. n = 12. About 50 species, southern Mexico to Paraguay, Antilles, equatorial Africa, Madagascar, Sri Lanka and southern India, Southeast Asia to northeastern Australia, New Caledonia; most speciose in Brazil.
 |
| Fig.40. Dilleniaceae. Tetracera boiviniana. A Flowering branch. B Stamens. C Gynoecium, vertical section. D Carpel, transverse section. E Fruit. F Arillate seed. (Gilg and Werdermann 1925) |
Section Tetracera with many-flowered thyrsoids with cincinnate partial inflorescences, small petals, and both simple and fasciculate trichomes; pantropical; sect. Akara Kubitzki with few(up to 12)-flowered thyrsoids, large, emarginate petals, and exclusively simple trichomes; OldWorld.
II. Subfam. Doliocarpoideae J.W. Horn (2005).
Smaller vessel elements with scalariform plates, commonly with fewer than 10 bars; rays extremely high, often > 10mm, some with uniseriate ends 1 or few cells high. Mature secondary stems producing successive cambia (except Curatella and perhaps also suffruticose Davilla spp.). Nodes (3)5(7)-lacunar; nodes bearing inflorescence axes, or in Davilla the primary inflorescence axis itself, producing subsequent flushes of inflorescences via proleptic proliferation buds. Inflorescences without prophylls, or prophylls irregularly produced in a few large-flowered species of Davilla; stigma conspicuously peltate, the margin annuliform and even; ovules 2 per carpel, typically 1 epitropous and 1 apotropous but the direction of ovule curvature variable among flowers of a given individual. When gynoecium apocarpous, fruits or partial fruits globose. Aril margin mostly entire; fleshy.
2. Davilla Vand.
Fig. 41
Davilla Vand., Fl. Lusit. Brasil.: 35 (1788); Kubitzki, Mitt. Bot. Staatssamml. München 9:1–105 (1971), rev.; Aymard in Fl. Venez. Guayana 4:671–685 (1998).
Scandent shrubs or lianas. Leaves sometimes with amplexicaul petiolar wings. Inflorescence a panicle with ultimate paraclades of triads or (less often) monads, never ramiflorous; sepals 5, free, the innermost 2 deeply concave, oppositely arranged, conspicuously larger than the ± flattened outer 3, and prominently accrescent; petals (3–)5; stamens 25–300, free; carpels 1–2(3). Fruit irregularly dehiscent, with very thin, papery walls, completely and indefinitely enclosed by the inner 2 accrescent sepals. Seeds partially or completely enclosed by a membranous, white aril. About 25 species, from southern Mexico to southern Brazil, Bolivia, and Paraguay; Antilles.
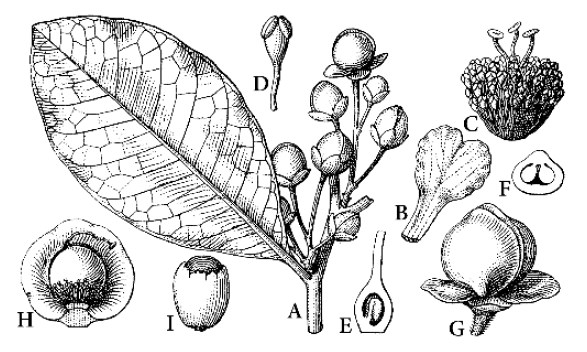 |
| Fig.41. Dilleniaceae. Davilla flexuosa. A Flowering branch. B Petal. C Androecium and gynoecium. D Stamen. E Carpel, vertical section. F same, transverse section. G Fruit enclosed by inner sepals. H Same after removal of one sepal. I Arillaate seed. (Gilg and Werdermann 1925) |
Section Homalochlaena Kubitzki, inner sepals with their margins pressed against each other; sect. Davilla, innermost sepalswith reflexedmargin and overlapped at margin by the second innermost sepal.
Other Doliocarpoideae:
Inflorescences exclusively axillary and (mostly) ramiflorous (possibly terminal in one Neodillenia sp.). Fruits with fleshy or leathery, typically brightly colored pericarps.
Curatella + Pinzona:
Leaf lamina decurrent on the petiole. Inflorescence a panicle. Gynoecium hemisyncarpous; carpels 2. Fruit a fleshy or leathery capsule, the stylodia crossing as the fruit matures on account of an unequal amount or rate of growth between the adaxial and abaxial sides of the carpels. Sepals reflexing after anthesis and hardly accrescent.
3. Curatella Loefl.
Fig. 38
Curatella Loefl., It. Hispan.: 260 (1758); Kubitzki, Mitt. Bot. Staatssamml. München 9:1–105 (1971), rev.
A small to medium-sized tree. Vestiture also including fasciculate trichomes. Leaves coarsely scabrous, owing to the presence of silicified epidermal cells and trichomes. Sepals (3)4(5), subequal, free; petals (3–)5, the median petal adaxial, or occasionally reduced or missing; stamens c. 80, free; carpels covered with fasciculate and hispid, simple trichomes. Pericarp outer surface green, inner surface scarlet. Seed completely enclosed by a membranous to slightly fleshy white aril. n = 13. One species, C. americana L., a tortuous savanna tree widespread fromsouthernMexico to southern Brazil and Bolivia, Antilles.
4. Pinzona Mart. & Zucc.
Pinzona Mart. & Zucc., Abh. Math.-Phys. Kl. Königl. Bayer. Akad. Wissensch. 1:371 (1832); Kubitzki, Mitt. Bot. Staatssamml. München 9:1–105 (1971), rev.; Aymard & Miller, Candollea 49:169–182 (1994).
A high-climbing liana, largely glabrous atmaturity except for inflorescence axes. Sepals 3–4, subequal, shortly basally connate; petals (2)3; stamens 25–35, free; carpels glabrous. Seed completely
enclosed within a fleshy orange aril. One species, P. coriacea Mart. & Zucc., throughout Central and South America from Belize to northeastern Brazil; Antilles.
5. Doliocarpus Rol.
Doliocarpus Rol., Kong. Svenska Vetensk. Acad. Handl. 17: 260 (1756); Kubitzki, Mitt. Bot. Staatssamml. München 9:1–105 (1971), rev.; Aymard & Miller, Candollea 49:169–182 (1994); Aymard, Anales Jard. Bot. Madrid 55:17–30 (1997);
Aymard in Fl. Venez. Guayana 4:671–685 (1998). Shrubs, mostly scandent, or lianas. Inflorescence a panicle, botryoid, or uncommonly a monad; the inflorescences at each node often appearing fasciculatedue tothe largenumberof accessorybudsproduced; sepals 3–6, free; petals 3–7; stamens c. 25–250, free; carpel 1(2). Fruit typically ripening red, baccate, dehiscing along both the dorsal and ventral side of the carpel (sometimes irregularly so), or indehiscent and berry-like. Seedswith a fleshy, entire, white aril. About 45 species, fromsouthernMexico and the Antilles to southern Brazil and Paraguay, with the center of species diversity in Brazil.
In sect. Calinea Eichl., the stamens are straight to slightly contorted in bud; in sect. Doliocarpus, the stamens are all reflexed in bud.
Genus dubium
5a. Neodillenia Aymard
Neodillenia Aymard, Harvard Pap. Bot. 10:121–131 (1997); Aymard in Fl. Venez. Guayana 4:671–685 (1998).
Lianas with successive cambia. Trichomes simple.
Inflorescences axillary (and sometimes also ramiflorous) and consisting of a solitary flower or botryoid, or terminal and consisting of a solitary flower or impoverished panicle (botryoid). Flowers large; sepals 3–6, unequal; corolla unknown; stamens 80–300, free; gynoecium apocarpous or merely shortly synovarious of (1)2–5 carpels; ovules (1)2 per carpel, said to be orthotropous (but, confusingly, also with a ventral raphe), though in N. peruviana Aymard they are clearly campylotropous, with one ovule epitropous and the other apotropous in each carpel. Seeds enclosed in a fleshy, entire, red aril. Fruit an aggregate of follicles, unknown in N. venezuelana Aymard. Three species, in the Amazonian regions of Colombia, Ecuador, Peru, and Venezuela.
Note that the stamens are entirely free (not shortly basally connate into a ring) and the ovules campylotropous (not orthotropous, cf. the orig. description). Neodillenia is clearly a member of subfamily Doliocarpoideae, as evidenced by its successive cambia, conspicuously peltate stigma with an even, annuliform margin, and ovular details. Neodillenia coussapoana and N. peruviana appear to be closest to large-flowered species of Doliocarpus sect. Doliocarpus, particularly D. grandiflorus and D. magnificus. The anther connectives of Neodillenia are of the same thickness as those of Doliocarpus magnificus and contain abundant raphid idioblasts, like Doliocarpus anthers. The only features that keep these two species separated from Doliocarpus are their red arils and gynoecia with typically 4–5 carpels. The monocarpellate N. venezuelana would be readily referable to Doliocarpus, except that it (apparently) has terminal inflorescences. Further work is needed to clarify both the structure and phylogenetic position of these plants.
Rest:
Vessel elements with exclusively scalariformperforation plates, or rarely alsowith fewsimple plates in a few xeromorphic Hibbertia species. Multiseriate wood rays with uniseriate ends always extended into long wings 4 or more cells in height (Kribs Type I). When inflorescences consisting of more than 1 flower, the β-prophyll often displaced onto the branch borne in its axil. Perianth nearly always 5-merous.Pollen grains uniformlywith simple, colpate apertures, typically 3-aperturate.
III. Subfam. Hibbertioideae J.W. Horn (2005).
6. Hibbertia Andrews
Figs. 42–44
Hibbertia Andrews, Bot. Rep.: t. 126 (1800); Bentham, Fl. Austral. 1:17–41 (1863); Hoogland, Fl. Males. I, 4:141–174 (1951); Stanley in Stanley & E.M. Ross, Fl. SE Queensland 1:185–189 (1983); Jessop in Jessop&Toelken, Fl. S.Australia 4th edn, 1:354–358 (1986); J.R. Wheeler in N.G. Marchant
et al., Fl. Perth Region 1:119–133 (1987); G.J. Harden & J. Evrett in G.J.Harden, Fl.NewSouthWales 1:293–303 (1990); Veillon in Fl. Nouv.-Caléd. Dépend. 16:3–86 (1990); Craven & Dunlop, Austral. Syst. Bot. 5:477–500 (1992), rev. of subg. Pachynema, p.p.;Wheeler in Fl. Kimberley Region 151–155 (1992); Toelken inWalsh & Entwisle, Fl. Victoria 3:300–313 (1996); Lewington & Cobb in Grieve, How to knowWestern Australian wildflowers, 2nd edn, 2:35–56 (1998); Murray in Fl. New South Wales suppl. 1:32–36 (2000); Wheeler in Fl. SouthWest: Bunbury-Augusta-Denmark 2:570–579 (2002); Wheeler, Nuytsia 15:311–320 (2004), key toW. Austral. spp.
Hemistema Thouars (1804).
Candollea Labill. (1806).
Pleurandra Labill. (1806).
Adrastaea DC. (1817).
Pachynema R. Br. ex DC. (1817).
Trisema Hook. f. (1857).
Shrubs, uncommonly small to medium-sized trees, or subshrubs, sometimes rhizomatous, with mostly or only cataphylls and with photosynthetic function transferred to the stems, rarely vines or lianas; nodes 1- or 3-lacunar. Leaves frequently ericoid, sometimes also with fasciculate, rarely peltate trichomes; venation semicraspedromous to brochidodromous, very rarely craspedromous, secondaries only occasionally parallel, and tertiaries rarely percurrent and never scalariform; areolation typically incomplete or lacking. Inflorescence commonly a monad, infrequently a cincinnus or thyrsoid with serial, cincinnate partial inflorescences, terminal and often also axillary, or infrequently apparently only axillary, uncommonly most of the plant body overtly inflorescence-like, and then a compound thyrsoid or panicle; sepals 5, unequal to ± equal, free or shortly fused; petals (3–)5, free; androecium of (1–)5–100(–300+) members, sometimes partly staminodial, basically polysymmetric or monosymmetric; polysymmetric androecia with the stamens all free, rarely all shortly basally connate, and ± evenly distributed around the carpels, or the stamens grouped into 3 or 5 distinct fascicles in which the stamen filaments may be all free, all shortly connate, or sometimes with 1 stamen free and the others in the fascicle connate; staminodes (when present) external to the fertile stamens, or (rarely) internal (subg. Pachynema); monosymmetric androecia always have the fertile stamens presented in the median plane of the flower and exclusively opposite the median petal, where they are free or fused into a single fascicle; staminodes, when present, external to and/or lateral to the fertile androecium, and uncommonly partly to entirely encircling the fertile stamens as a unit; anthers dehiscence via longitudinal slits or, sometimes, via apical pores; carpels 1–5(–10), glabrous or pubescent; ovules 1–25 per carpel. Seeds 1–8 per follicle; aril subfleshy or pulpy and oily or waxy, whitish, subentire, or rarely red and fleshy. About 225 species, from Madagascar (1 sp.) to Fiji (1 sp.), c. 200 species in Australia incl. Tasmania, 24 species in New Caledonia, 2 species in New Guinea. The division into four subgenera, presented here, is strongly supported by both molecular and morphological data (Horn 2005; J.W. Horn, unpubl. data).
6a. Hibbertia subg. Pachynema (R. Br. ex DC.) J.W. Horn (2005).
Fig. 42
Subshrubs, sometimes rhizomatous; true leaves (if present) confined to basalmost nodes of a shoot, with craspedromous venation (the only instance of thiswithin the genus); aerial axes green and photosynthetic, often caespitose and sometimes dimorphic, provided withmostly or (typically) only cataphylls. Inflorescence (which constitutes nearly the whole of the shoot system) a compound thyrsoidor panicle, commonly with serial branches or flowers; and roecium typically bicyclic, with an outer whorl of 7(–10) fertilestamensandaninnerwhorl of 2 stamens or staminodes in the transverse plane of the flower; infrequently the androecium unicyclic and consistingof4–5 stamens, apparentlyby reduction; carpels 2, ovules 2 per carpel. n = 12; 2n = 30. Nine species, including all species recognized by Craven and Dunlop (1992) in Pachynema, plus Hibbertia conspicua (Harv.) Gilg and H. goyderi F. Muell., in Australia, mostly confined to the Northern Territory (especially the Arnhem Plateau) and adjacent regions of the Kimberley of Western Australia, and in the Geraldton sandplains of the Southwest Botanical Province of Western Australia.
 |
| Fig.42. Dilleniaceae. A Hibbertia (subg. Pachynema) dilatatum, flowering shoot. C-F Hibbertia (sug.Pachynema) complanatum. C Flower bud. D Same seen from below. E Flower. F Androecium and gynoecium, vertical section. ( Gilg and Werdermann 1925) |
6b. Hibbertia subg. Hemistema (Thouars) J.W. Horn (2005).
Fig. 43
Small shrubs to medium-sized trees; mature axes non-photosynthetic. Leaves moderately large and broad to (frequently) small and ericoid; vestiture frequently also including fasciculate trichomes (sometimes only), or infrequently including multiradiate to peltate trichomes. Inflorescence commonly a terminal monad (often also axillary, or terminal on sometimes exclusively axillary short shoots), less often a terminal (and also often axillary) cincinnus or thyrsoid; androecium monosymmetric or less frequently polysymmetric and then the stamens not aggregated into either clearly defined cycles or fascicles; carpels basi cally and commonly 2, uncommonly 3–5 due to a secondary increase in number, rarely 1; ovules 1–25 per carpel. n = 9; 2n = 26, 36. Estimated 160 species, including all species with monosymmetric androecia, all New Caledonian species, all species
included in, or attributable to, Gilg and Werderman’s (1925) section Cyclandra series Ochrolasiae, Tomentosae, and Vestitae, plus Hibbertia arcuata J.R. Wheeler, H. graniticola J.R. Wheeler, and the H. exasperata group (Wheeler 2004b), which has been previously associated with section Candollea. Distribution equivalent to that of the whole genus.
 |
| Fig.43. Dilleniaceae. Hibbertia (subg. Hemistema). baudouini. A Flowering branch. B Androecium spread out and gynoecium. C Stamen. D Carpel, vertical section. E Carpel, transverse section. F Arillate seed. (Gilg and Werdermann 1025) |
6c. Hibbertia subg. Adrastaea (R. Br. ex DC.) J.W. Horn (2005).
A wiry shrub, becoming scandent with age; mature axes non-photosynthetic. Leaves laminar, small and linear. Inflorescence a terminalmonad; flowers initially produced on axillary short shoots, later on sympodial long shoots consisting mostly of 1-flowered, 2-prophyllate modules; stamens 10 in 2 cycles, obdiplostemonous; carpels 2, ovule 1 per carpel. Only one species, H. salicifolia (DC.) F. Muell., in wallum heath near the coast of temperate to subtropical eastern Australia, from just south of Sydney north to the Bundaberg area in southeastern Queensland.
 |
| Fig.44. Dilleniaceae. Hibbertia (subg. Hibbertia) scandens. A Flowering branch. B Flower bud. C Androecium and gynoecium, vertical section. D Stamen. E Gynoecium, tansverse section. F Fruit. G Seed enclosed by aril. H Same, vertical section. (Gilg and Werdermann 1925) |
6d. Hibbertia subg. Hibbertia
Fig. 44
Shrubs or, uncommonly, vines or lianas; mature axes non-photosynthetic. Leaves laminar, or ericoid in the Hibbertia hemignosta species complex within sect. Candollea. Stamens not aggregated into obvious cycles, or rarely 1 cycle in the few cases where there are 3 stamens, but sometimes groupedinto 3 or 5 alternipetalous fascicles; carpels (1–)3(4)5(–15), fundamentally and commonly 3 or 5; ovules 1–10 per carpel. n = 4, 5, 8, 16, 32, 64; 2n = 20. Estimated 80–90 species, containing all species included in or attributable to the series of Gilg andWerdermann’s (1925) sect. Cyclandra not accounted for above, plus those in or attributable to their sect. Candollea (except the H. exasperata group, see above). Occurring throughoutAustralia, but particularly concentrated in the southeast and southwest, New Guinea.
IV. Subfam. Dillenioideae Burnett (1835)(‘Dillenidae’).
Nodes multilacunar (7–27), or probably secondarily trilacunar in the Sri Lankan species of Acrotrema. Leaves with persistent or deciduous, amplexicaul petiolar wings (perhaps secondarily lost in one Dillenia clade).
Schumacheria + Didesmandra:
Shrubs to small, spindly trees; androecium monosymmetric; pollen commonly 4-aperturate; ovule 1 per carpel, campylotropous, apotropous; vascularized by a massive bundle of traces that remain in the floral receptacle above the point where the ventral carpel bundles depart.
7. Schumacheria Vahl
Fig. 45
Schumacheria Vahl, Skr. Naturhist.-Selsk. 6:122 (1810); Wadhwa, Rev. Handb. Fl. Ceylon 10:109–135 (1996).
Inflorescence both terminal and axillary, a thyrsoid with a paniculate arrangement of cincinnate partial inflorescences in S. castaneifolia, or consisting of axillary cincinni in the other species. Sepals 5; petals 3–5; androecium of c. 20–35 stamens, grouped in a single fascicle located opposite the abaxial-median petal; stamen filaments nearly fully connate, forming a liguliform column; anthers with a vestiture of simple trichomes and with a short, mucronate, apical appendage, dehiscing by 2 apical pores; carpels (2)3. Seed with small, membranous aril. Three species, endemic to Sri Lanka.
 |
| Fig.45. Dilleniaceae. Schumacheria castaneifolia. A Flowering branch. B Flowe bud. C Androecium and gynoecium. D Anther. E Carpel, vertical section. F Carpel, transverse section. G Seed. (Gilg and Werdermann 1925) |
8. Didesmandra Stapf
Didesmandra Stapf in Hooker’s Ic. Pl.: t. 2646 (1900);Hoogland, Fl. Males. I, 4:152, Fig. 7 (1951).
Vestiture also including fasciculate trichomes. Inflorescence a terminal (lessoftenalsoaxillary) thyrsoid with cincinnate partial inflorescences, at least some of which represent supernumerary branches; sepals 5; petals 5; stamens grouped in 2 fascicles presented in the median plane of the flower, opposite the median petal with the filaments in each fascicle connate to form a ± cylindrical staminal column, the two stamen fascicles very shortly connate with one another at their base; each of the two stamen fascicles consists of 1 larger stamen with a conspicuously uncinate anther with dehiscence by longitudinal slits and 4 smaller stamenswith unbent anthers that dehisce by apical pores; carpels 2, presented in the transverse plane of the flower.
Fruit an aggregate of nutlets. Seed with a membranous aril. One species, D. aspera Stapf, endemic
to Sarawak, Borneo.
Acrotrema + Dillenia:
Gynoeciumhemisyncarpous.Ovules 2 ormore per carpel.
9. Acrotrema Jack
Fig. 46
Acrotrema Jack, Mal. Misc. 1, 5:36 (1820); Hoogland, Fl. Males. I, 4:141–174 (1951); Hoogland, Fl. Thailand 2, 2:95–108 (1972); Wadhwa, Rev. Handb. Fl. Ceylon 10:109–135 (1996).
Rhizomatous herbs with leaves in a basal rosette or terminal on a very short, erect stem. Leaves simple, pinnatisect, or pinnate, sometimes variegated, the base sometimes auriculate; vestiture of simple, unicellular or multicellular trichomes. Inflorescence terminal, or sometimes also axillary, a raceme, or theflower solitary; sepals 5; petals 5; androeciumof 15–50 free stamens, usually grouped in 3 fascicles positioned alternate with the carpels, or stamens evenly distributed around the gynoecium; anthers short and dehiscing via longitudinal slits, or long, linear, and dehiscing via 2 apicalpores; carpels (2)3; ovules 2–6(–20) per carpel. Fruit an aggregate of basally coherent follicles, dehiscing irregularly at maturity. Seed with a white and membranaceous aril. n = 28 (Mathew 1972). About 9 species, c. 7 in Sri Lanka, 1 in the Western Ghats of India, and 1 in southernMyanmar, southern Thailand,Malay Peninsula, and northern Sumatra.
Further work is needed to determine whether this genus is derived within, or is sister to, Dillenia.
 |
| Fig.46. Dilleniaceae. A Acrotrema thwaitesii. B Acrotrema lanceolatum. (Gilg and Werdermann 1925) |
10. Dillenia L.
Figs. 47, 48
Dillenia L., Sp.
Pl. 1:535 (1753); Hoogland, Fl. Males. I, 4:141–174 (1951), in Blumea 7:1–145 (1952), rev., ibid. 9:577–589 (1959), and Fl. Thailand 2, 2:95–108 (1972).
Trees, rarely shrubs, mostly evergreen, rarely deciduous. Leaves with or without persistent or deciduous amplexicaul petiolar wings. Inflorescences 1–10(–30)-flowered, a cincinnus, thyrsoid with (apparently) cincinnate partial inflorescences, panicle (often depauperate), or monad, terminal or rarely axillary; flowers infrequently borne on cataphyll-bearing short shoots; sepals (4)5(–18), unequal to ± equal; petals (0, 4)5(–7); androecium of (60–)100–700(–900) members, sometimes partly staminodial; fertile stamens often distinctly heterantherous (the inner group of stamens bearing anthers overtly longer than those of the outer group); anthers dehiscence frequently via 2 apical or subapical pores, less frequently via 2 longitudinal slits; staminodes, when present, typically external to the fertile androecium, rarely internal; gynoecium typically (always?) hemisyncarpous; carpels (4)5–15(–20), forming one whorl on a conical, centrally protruding part of the receptacle; ovules 5–80 per carpel, borne in 2(4) vertical rows on submarginal placentae. Fruit an aggregate of basally coherent follicles, or the fruit indehiscent and completely enclosed by the fleshy, accrescent calyx. Seeds with a white or red, fleshy aril, or the aril vestigial in some species with indehiscent fruits. n = 13, 16, 24, 27. About 65 species, from Madagascar and the Seychelles to Fiji, c. 45 species in Malesia, 1 in Australia.
The non-amplexicaul species group, recognized by Hoogland (1952), is monophyletic (Horn 2005), but some of the amplexicaul species group (e.g., D. triquetra of Madagascar and Sri Lanka; possibly D. ferruginea of the Seychelles) may be most closely related to Acrotrema.
 |
| Fig.47. Dilleniaceae. Dillenia excelsa. A Flowering branch. B Flower. C Dehiscing fruit. (Koorders and Valeton 1913) |
 |
| Fig.48. Dilleniaceae. Dillenia indica. A Flowering branch. B Androecium and gynoecium, vertical section. C Half gynoecium, transverse section. D Outer and inner stamen. E Fruit enclosed by fleshy sepals. F Same, vertical section. G Seed. H Same, vertical section. (Gilg and Werdermann 1925) |
Selected Bibliography
APG II 2003. See general references.
Ashton, P.S., Gunatilleke, C.V.S. 1987. New light on the plant
geography of Ceylon. I. Historical plant geography. J.
Biogeogr. 14:249–285.
Aymard, C., G.A. 1997. Dilleniaceae novae Neotropicae IX:
Neodillenia, a new genus fromthe Amazon basin.Harvard
Pap. Bot. 10:121–131.
Aymard, C., G.A. 2002. Davilla papyracea (Dilleniaceae),
a new species from Brazil. Kew Bull. 57:487–490.
Baillon, H.E. 1865. Remarques sur les Dilléniacées. Adansonia
6:255–281.
Baretta-Kuipers, T. 1972. Some remarks on the wood structure
of Pinzona and allied genera of the subfamily
Tetraceroideae (Dilleniaceae). Acta Bot. Neerl. 21:573–
577.
Berg, R.Y. 1975. Myrmecochorous plants in Australia and
their dispersal by ants. Austral. J. Bot. 23:475–508.
Bernhardt, P. 1984. The pollination biology of Hibbertia
stricta (Dilleniaceae). Pl. Syst. Evol. 147:267–277.
Bernhardt, P. 1986. Bee-pollination in Hibbertia fasciculata
(Dilleniaceae). Pl. Syst. Evol. 152:231–241.
Bernhardt, P. 1996. Anther adaptation in animal pollination.
In: D’Arcy, W.G., Keating, R.C. (eds) The anther:
form, function,andphylogeny.Cambridge:Cambridge
University Press, pp. 192–220.
Conti, E. et al. 2002. See general references.
Corner, E.J.H. 1946. Centrifugal stamens. J. Arnold Arb.
27:423–437.
Corner, E.J.H. 1976. See general references.
Corner, E.J.H. 1978. The inflorescence of Dillenia. Notes
Roy. Bot. Gard. Edinburgh 36:341–353.
Craven, L.A., Dunlop, C.R. 1992. A taxonomic revision of
Pachynema (Dilleniaceae). Austral. Syst. Bot. 5:477–
500.
Croat, T. 1978. Flora of Barro Colorado Island. Stanford:
Stanford University Press.
Cronquist, A. 1981. See general references.
Cronquist, A. 1988. The evolution and classification of flowering
plants, 2nd edn. Bronx: New York Botanical Garden.
Cuénoud, P., Savolainen, V., Chatrou, L.W., Powell, M.,
Grayer, R.J., Chase, M.W. 2002. Molecular phylogenetics
of Caryophyllales based on 18S rDNA and plastid
rbcL, atpB, and matK DNA sequences. Amer. J. Bot.
89:132–144.
Dahlgren, R.M.T. 1983. General aspects of angiospermevolution
and macrosystematics. Nordic J. Bot. 3:119–
149.
de Candolle, A.P. 1824. Prodromus systematis naturalis
regni vegetabilis, I. Paris: Treuttel et Würtz.
Dickison, W.C. 1967a. Comparative morphological studies
in Dilleniaceae, I.Wood anatomy. J. Arnold Arb. 48:1–
29.
Dickison, W.C. 1967b. Comparative morphological studies
in Dilleniaceae, II. The pollen. J. Arnold Arb. 48:231–
240.
Dickison, W.C. 1968. Comparative morphological studies
in Dilleniaceae, III. The carpels. J. Arnold Arb. 49:317–
333.
Dickison, W.C. 1969. Comparative morphological studies
in Dilleniaceae, IV. Anatomy of the node and vascularization
of the leaf. J. Arnold Arb. 50:384–410.
Dickison, W.C. 1970a. Comparative morphological studies
inDilleniaceae, V. Leaf anatomy. J. Arnold Arb. 51:89–
113.
Dickison, W.C. 1970b. Comparative morphological studies
in Dilleniaceae, VI. Stamens and young stem. J. Arnold
Arb. 51:403–422.
Dickison, W.C. 1971. Comparative morphological studies
in Dilleniaceae, VII. Additional notes on Acrotrema. J.
Arnold Arb. 52:319–333.
Dickison, W.C. 1979. A note on the wood anatomy of Dillenia
(Dilleniaceae). IAWA Bull. 2/3:57–60.
Dickison, W.C. 1984. On the occurrence of silica grains in
the woods of Hibbertia (Dilleniaceae). IAWA Bull. II,
5:341–343.
Dickison, W.C., Rury, P.M., Stebbins, G.L. 1978. Xylem
anatomy of Hibbertia (Dilleniaceae) in relation to
ecology and evolution. J. Arnold Arb. 59:32–49.
Dickison,W.C.,Nowicke, J.W., Skvarla, J.J. 1982. Pollenmorphology
of the Dilleniaceae and Actinidiaceae. Amer.
J. Bot. 69:1055–1073.
Dyer, A.G. 1996. Reflection of near-ultraviolet radiation
from flowers of Australian native plants. Austral. J.
Bot. 44:473–488.
Eichler, A.W. 1878. Blüthendiagramme, II. Leipzig: W. Engelmann.
Endress, P.K. 1997. Relationships between floral organization,
architecture, and pollination mode in Dillenia
(Dilleniaceae). Pl. Syst. Evol. 206:99–118.
George, A.S. 2002. The south-western Australian flora in
autumn: 2001 PresidentialAddress. J.Roy. Soc.W.Australia
85:1–15.
Gilg, E., Werdermann, E. 1925. Dilleniaceae. In: Engler, A.,
Prantl, K., Die natürlichen Pflanzenfamilien, ed. 2, 21.
Leipzig:W. Engelmann, pp. 7–36.
Gottsberger, G. 1977. Some aspects of beetle pollination in
the evolution of flowering plants. Pl. Syst. Evol., suppl.
1:211–226.
Gurni, A.A., Kubitzki, K. 1981. Flavonoid chemistry and
systematics of the Dilleniaceae. Biochem. Syst. Ecol.
9:109–114.
Hegnauer, R. 1966. Chemotaxonomie der Pflanzen, 4, pp.
19–23. Basel: Birkhäuser.
Hickey, L.J., Wolfe, J.A. 1975. The bases of angiosperm
phylogeny: vegetativemorphology.Ann.Missouri Bot.
Gard. 62:538–589.
Hoogland, R.D. 1952. A revision of the genus Dillenia.
Blumea 7:1–145.
Hoogland, R.D. 1953. The genus Tetracera (Dilleniaceae) in
the eastern OldWorld. Reinwardtia 2:185–225.
Hoogland, R.D. 1959. Additional notes on Dilleniaceae 1–9.
Blumea 9:577–589.
Horn, J.W. 2005. The phylogenetics and structural botany
of Dilleniaceae and Hibbertia Andrews. Ph.D. Thesis,
Duke University, Durham, NC, 171 p.
Hutchinson, J. 1964. The genera of flowering plants, 1. Oxford:
Clarendon.
Imaichi, R., Kato, M. 1996. A scanning electron microscopic
study of ovule development in Dillenia suffruticosa
(Dilleniaceae). Phytomorphology 46:45–51.
Ioffe, M.D., Zhukova, G.Y. 1974. Modification of zygote cell
wall during its development in Dillenia Dilleniaceae.
Bot. Zhurn. (Moscow & Leningrad) 59:1409–1416.
Keighery, G.J. 1975. Pollination of Hibbertia hypericoides
(Dilleniaceae) and its evolutionary significance. J. Nat.
Hist. 9:681–684.
Keighery, G.J. 1991. Pollination of Hibbertia conspicua (Dilleniaceae).
W. Austral. Naturalist 18:163–165.
Koorders, S.H., Valeton, T. 1913. Atlas der Baumarten von
Java, 1. Leiden: P.W.M. Trap.
Kubitzki, K. 1968. Flavonoide und Systematik der Dilleniaceen.
Ber. Deutsch. Bot. Gesell. 81:238–251.
Kubitzki, K. 1970. Die Gattung Tetracera (Dilleniaceae).
Mitt. Bot. Staatssamml. München 8:1–98.
Kubitzki, K. 1971. Doliocarpus, Davilla und verwandte
Gattungen (Dilleniaceae). Mitt. Bot. Staatssamml.
München 9:1–105.
Lakshmanan, J.D.E., Lakshmanan, K.K. 1984. Further contributions
to the embryology of Dillenia suffruticosa
(Griff.) Martelli. J. Indian Bot. Soc. 63:353–359.
Mathew, P.M. 1972.Cytology ofAcrotrema.Curr. Sci. 41:751.
Momose, K., Yumoto, T., Nagamitsu, T., Kato, M.,
Nagamasu, H., Sakai, S., Harrison, R.D., Itioka, T.,
Hamid, A.A., Inoue, T. 1998. Pollination biology in
a lowland dipterocarp forest in Sarawak, Malaysia.
I. Characteristics of the plant-pollinator community
in a lowland dipterocarp forest. Amer. J. Bot.
85:1477–1501.
Ozenda, P. 1949. Recherches sur les dicotylédons apocarpiques.
Publications des laboratoires de l’École Normale
Supérieure, Série Biologie, fasc. 2. Paris: Jouve.
Paetow, W. 1931. Embryologische Untersuchungen an Taccaceen,
Meliaceen und Dilleniaceen. Planta 14:441–
470.
Parmentier, M.P. 1895. Contribution à l’étude de la famille
des Dilléniacées. C. R. Assoc. Franç. Avancem. Sci. pt.
2:626–630.
Pavanasasivam, G., Sultanbawa, M.U.S. 1974. Betulinic acid
in the Dilleniaceae and a review of its natural distribution.
Phytochemisty 13:2002–2006.
Prance, G.T. 2003. Rhabdodendraceae. In: Kubitzki, K.,
Bayer, C. (eds) The Families and Genera of Vascular
Plants, 5. Flowering plants, dicotyledons: Malvales,
Capparales and non-betalain Caryophyllales. Berlin
Heidelberg New York: Springer, pp. 339–341.
Rao, A.N. 1957. A contribution to the embryology of Dilleniaceae.
Proc. Iowa Acad. Sci. 64:172–176.
Rao, T.A., Das, S. 1979. Comparative typology and taxonomic
value of foliar sclereids in Hibbertia Andr. (Dilleniaceae).
Proc. Indian Acad. Sci., B 88:161–174.
Rury, P.M., Dickison, W.C. 1977. Leaf venation patterns
of the genus Hibbertia (Dilleniaceae). J. Arnold Arb.
58:209–256.
Sastri, R.L.N. 1958. Floral morphology and embryology of
some Dilleniaceae. Bot. Notiser 111:495–511.
Schatral, A. 1995. The structure of the seed in some Western
Australian species of the genus Hibbertia (Dilleniaceae).
Bot. J. Linn. Soc. 119:257–263.
Schatral, A. 1996. Dormancy in seeds of Hibbertia hypericoides
(Dilleniaceae). Austral. J. Bot. 44:213–222.
Schatral, A., Kailis, S.G., Fox, J.E.D. 1994. Seed dispersal of
Hibbertia hypericoides (Dilleniaceae) by ants. J. Roy.
Soc.W. Australia 77:81–85.
Soltis, D.E. et al. 2003. See general references.
Stebbins, G.L., Hoogland, R.D. 1976. Species diversity,
ecology and evolution in a primitive angiosperm
genus: Hibbertia (Dilleniaceae). Pl. Syst. Evol.
125:139–154.
Steppuhn, H. 1895. Beiträge zur vergleichenden Anatomie
der Dilleniaceen. Bot. Centralbl. 62:337–342, 369–378,
401–413.
Swamy, G.L., Periasamy, K. 1955. Contributions to the embryology
of Acrotrema arnottianum. Phytomorphology
5:301–314.
Takhtajan, A. 1997. See general references.
Thorne, R.F. 2000. The classification and geography of flowering
plants: dicotyledons of the class Angiospermae.
Bot. Rev. (Lancaster) 66:441–647.
Toelken, H.R. 1998. Notes on Hibbertia (Dilleniaceae): 2.
The H. aspera–empetrifolia complex. J. Adelaide Bot.
Gard. 18:107–160.
Toelken, H.R. 2000. Notes on Hibbertia (Dilleniaceae): 3. H.
sericea and associated species. J. Adelaide Bot. Gard.
19:1–54.
Tucker, S.C., Bernhardt, P. 2000. Floral ontogeny, pattern
formation, and evolution in Hibbertia and Adrastaea
(Dilleniaceae). Amer. J. Bot. 87:1915–1936.
Veillon, J.-M. 1990. Dilleniaceae. In:Morat, P.,MacKee, H.S.
(eds) Flore de la Nouvelle-Calédonie et Dépendances,
16, pp. 3–86. Paris: Muséum National d’Histoire Naturelle.
Vyshenskaya, T.D., Oganezova, G.G. 1991. Dilleniaceae.
In: Takhtajan, A. (ed.) Anatomia seminum comparativa
(in Russian), 3, pp. 163–171. Leningrad:
Nauka.
Wagner, R. 1906a. Über den Bau der Rispen des Trisema
Wagapii Vieill. Sitzungsber. Kaiserl. Akad. Wissensch.,
Math.-Naturwissensch. Kl., Abt. 1, 115:857–
880.
Wagner, R. 1906b. Untersuchungen über den morphologischen
Aufbau der Gattung Pachynema R. Br. Sitzungsber.
Kaiserl.Akad.Wissensch.,Math.-Naturwissensch.
Kl., Abt. 1, 115:1039–1080.
Wheeler, J.R. 1984. New species of Hibbertia (Dilleniaceae)
fromthe northernwheatbelt area ofWesternAustralia.
Nuytsia 9:427–437.
Wheeler, J.R. 2002. Miscellaneous new species of Hibbertia
(Dilleniaceae) from the wheatbelt and pastoral areas
ofWestern Australia. Nuytsia 15:139–152.
Wheeler, J.R. 2004a. A review of Hibbertia hemignosta and
its allies (Dilleniaceae) fromWestern Australia. Nuytsia
15:277–298.
Wheeler, J.R. 2004b. An interim key to the Western
Australian species of Hibbertia (Dilleniaceae). Nuytsia
15:311–320.
Wilson, C.L. 1937. The phylogeny of the stamen. Amer. J.
Bot. 24:686–699.
Wilson, C.L. 1965. The floral anatomy of the Dilleniaceae. I.
Hibbertia Andr. Phytomorphology 15:248–274.
Wilson, C.L. 1973. The floral anatomy of theDilleniaceae. II.
Generaother than Hibbertia.Phytomorphology 23:25–
42.












